
Noise-driven Fate Selection

Since the 1960s, it had been hypothesized that organisms might harness intrinsic variability to enable bet-hedging decisions between alternate developmental fates (Cohen, 1966), in much the same way that financial houses diversify their assets to minimize risk in volatile markets.
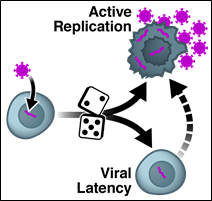
Our studies demonstrated, for the first time, that stochastic 'noise' in gene expression—Brownian fluctuations arising from diffusion-limited reactions—is harnessed to drive a biological fate decision (Weinberger et al., 2005). By pioneering the study of HIV’s decision circuit, we demonstrated that gene-expression noise enables HIV's to select between active replication and a long-lived dormant state (proviral latency), which is the chief barrier to achieving an HIV cure.
Noise in gene expression is now understood to drive fate selection in systems ranging from bacteria to cancer (Balaszi et al. Cell 2012).
For review: Singh and Weinberger. Curr Opinion 2009
Papers of note: Weinberger**, et al. Cell 2005 (linked above) and Weinberger**, Dar, Simpson. Nature Genetics 2008